Fructose is metabolized differently than glucose with up to 75% of orally absorbed fructose being metabolized by the liver. Moreover, the key enzyme of fructose metabolism, fructokinase is not feedback regulated by fructose metabolites. Thus, all of the orally absorbed fructose is phosphorylated by fructokinase. Unregulated AbMole Mepiroxol metabolism of fructose causing ATP depletion, oxidative stress and decrease in nitric oxide production has been proposed as one of the mechanisms of fructose-induced increase in blood pressure. Both glucose and fructose are precursors of MG formation and MG is also a well-established trigger for increased oxidative stress through multiple pathways. MG is also a major precursor of the formation of AGEs. The aorta and the kidney of fructose treated rats had significantly elevated MG AbMole 12-O-Tiglylphorbol-13-isobutyrate levels which implicates MG as a factor in hypertension development. The kidney had increased protein expression of renin, which in turn would produce more Ang II and its multiple effects. The increased Ang II may be responsible for the increased a1D receptor expression. The increased Ang II, AT1 and a1D receptors in the aorta signify increased vascular tone. We were not able to harvest enough tissue from the mesenteric artery for western blot and PCR analysis. GSH levels were reduced in the aorta and the kidney of fructose treated rats. GSH plays a central role in the degradation of MG by binding MG and making it available to the glyoxalase enzymes. A reduction in GSH would decrease MG degradation, increase its levels, and set up a vicious cycle. We addressed the possibility that MG was increasing oxidative stress, which has been implicated in the pathogenesis of hypertension. The effect of MG on RAGE has not been reported before. RAGE is currently under intense investigation as a target to prevent diabetic complications. The activation of RAGE by AGEs has been reported to increase two key transcription factors, NF-kB and early growth response-1, and cause oxidative stress. We observed increased RAGE and NF-kB in the aorta and the kidney of fructose treated rats. These results suggest that fructose treatment increases MG levels, which in turn can activate RAGE, which then increases NFkB and oxidative stress. The increased oxidative stress increases expression of Ang II, AT1 and a1D receptors. The increased Ang II in turn can increase NF-kB and oxidative stress and set up a vicious cycle. Our results suggest  a possible sequence of molecular events resulting from elevated endogenous MG levels which have been reported in high carbohydrate diet fed animals and in diabetic patients. Metformin is used clinically as an oral anti-diabetic drug in patients with type 2 diabetes. Metformin has multiple effects in vivo. For example, it inhibits mitochondrial respiration and gluconeogenesis in the liver, activates AMP-activated protein kinase, increases insulin sensitivity, antagonizes the action of glucagon and increases fatty acid oxidation. These in vivo antidiabetic actions of metformin can make interpretation of results difficult, making it a less than ideal experimental MG scavenger. Even though the MG scavenging ability of metformin has been studied in specific experiments and reported before, we tested the MG scavenging ability of metformin in cultured vascular smooth muscle cells treated with MG, where multiple in vivo, especially hepatic, actions of metformin do not come into play. We also performed an in vitro MG scavenging assay for metformin. In cultured vascular smooth muscle cells metformin attenuated the increased expression of a1D and AT1 receptors, and Ang II.
a possible sequence of molecular events resulting from elevated endogenous MG levels which have been reported in high carbohydrate diet fed animals and in diabetic patients. Metformin is used clinically as an oral anti-diabetic drug in patients with type 2 diabetes. Metformin has multiple effects in vivo. For example, it inhibits mitochondrial respiration and gluconeogenesis in the liver, activates AMP-activated protein kinase, increases insulin sensitivity, antagonizes the action of glucagon and increases fatty acid oxidation. These in vivo antidiabetic actions of metformin can make interpretation of results difficult, making it a less than ideal experimental MG scavenger. Even though the MG scavenging ability of metformin has been studied in specific experiments and reported before, we tested the MG scavenging ability of metformin in cultured vascular smooth muscle cells treated with MG, where multiple in vivo, especially hepatic, actions of metformin do not come into play. We also performed an in vitro MG scavenging assay for metformin. In cultured vascular smooth muscle cells metformin attenuated the increased expression of a1D and AT1 receptors, and Ang II.
These results support a direct effect of MG on these RAAS mediators and the MG scavenging action of metformin
Leave a reply
 of secondary structure populations in disordered proteins, to assess directly the conformation of aSyn within living cells.
of secondary structure populations in disordered proteins, to assess directly the conformation of aSyn within living cells. way as WNN, giving the Const-GIP method. With this method all possible interactions for drug/targets with only one interaction will be ranked before interactions with drugs/targets that also have other interactions. Essentially, for such interactions the method only has to do half the work, since the fact that the drug/target is correct can be known with certainty. In real world situations there are also drug compounds that interact with none of the target under consideration, and vice versa, which would invalidate the ConstGIP method. This resulted in a similarity matrix for the denoted by Sc, which represents the chemical space. Amino acid sequences of the target proteins were obtained from the KEGG GENES database. Sequence similarity between proteins was computed using a normalized version of Smith-Waterman score, resulting in a similarity matrix denoted Sg, which represents the genomic space. We could test the prediction capability of the proposed methods on unknown drug-target interactions of the given network using the procedure adopted in. Therein, the complete interaction network for each dataset is used as training data, and the predictions on non-interacting pairs in the training set are ranked with respect to their interaction scores. However, since each drug compound or target in the training set has at least one interaction, we do not need to use WNN and the results are those of GIP. We report the top five predicted interactions for each dataset in Table 3. In general, these results indicate that cross validation should be applied and interpreted with care. Note that the cross validation procedure used in the comparison with KBMF2K is also positively biased, since we know that each ‘new’ drug compound has at least one interaction, but there the bias is much smaller. In this work, we proposed a simple yet effective procedure to predict interaction profiles for unknown drug compounds and show how it can be directly integrated into a recent machine learning algorithm for the in-silico prediction of drug-target interactions. The novelty of our approach comes in the use of a weighted nearest neighbor procedure for inferring a profile for a drug compound by using interaction profiles of the compounds in the training data, where each profile is weighted using information about chemical similarity between drug compounds integrated with a simple decay scheme.
way as WNN, giving the Const-GIP method. With this method all possible interactions for drug/targets with only one interaction will be ranked before interactions with drugs/targets that also have other interactions. Essentially, for such interactions the method only has to do half the work, since the fact that the drug/target is correct can be known with certainty. In real world situations there are also drug compounds that interact with none of the target under consideration, and vice versa, which would invalidate the ConstGIP method. This resulted in a similarity matrix for the denoted by Sc, which represents the chemical space. Amino acid sequences of the target proteins were obtained from the KEGG GENES database. Sequence similarity between proteins was computed using a normalized version of Smith-Waterman score, resulting in a similarity matrix denoted Sg, which represents the genomic space. We could test the prediction capability of the proposed methods on unknown drug-target interactions of the given network using the procedure adopted in. Therein, the complete interaction network for each dataset is used as training data, and the predictions on non-interacting pairs in the training set are ranked with respect to their interaction scores. However, since each drug compound or target in the training set has at least one interaction, we do not need to use WNN and the results are those of GIP. We report the top five predicted interactions for each dataset in Table 3. In general, these results indicate that cross validation should be applied and interpreted with care. Note that the cross validation procedure used in the comparison with KBMF2K is also positively biased, since we know that each ‘new’ drug compound has at least one interaction, but there the bias is much smaller. In this work, we proposed a simple yet effective procedure to predict interaction profiles for unknown drug compounds and show how it can be directly integrated into a recent machine learning algorithm for the in-silico prediction of drug-target interactions. The novelty of our approach comes in the use of a weighted nearest neighbor procedure for inferring a profile for a drug compound by using interaction profiles of the compounds in the training data, where each profile is weighted using information about chemical similarity between drug compounds integrated with a simple decay scheme. research. The general approach developed below is to construct a hypothetical trial that embodies the known characteristics of the treatment and pharmacogenomic marker.
research. The general approach developed below is to construct a hypothetical trial that embodies the known characteristics of the treatment and pharmacogenomic marker.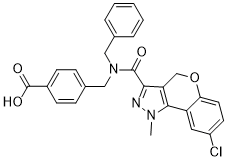 LV EF from baseline to 4 months was less pronounced in patients with MVO, as compared to those without. Thus, the presence of MVO, as demonstrated by a CMR study during the first few days after STEMI, is a marker of an unfavorable recovery over time of jeopardized myocardium. MVO was also associated with a large final infarct size, and this association was still highly significant after adjustment for other markers of outcome, including myocardium at risk. Of note, none of our patients had collateral flow to IRA and, except for 3 patients with TIMI 2-flow, all patients in our study had normalization of macrovascular flow after PCI. Our results confirm and extend previous reports on the association between MVO and variables of LV function after PCI-treated STEMI. In patients with presence of MVO, we also found change in LV ESV between baseline and 4 months than in patients without MVO. However, in contrast to other reports, after adjustment for confounders, the change in LV EDV was not significantly different in the 2 groups. Other baseline variables, including myocardium at risk, were of importance for the remodeling process, as expressed by a significant association with LVEDV. This observation corresponds with the results of Lund and coworkers, concluding that infarct size measured by early CMR is a stronger predictor of remodeling than MVO. Some investigators have calculated the size and extent of MVO and related this to LV outcome variables. However, according to Nijveldt and co-workers, it is the presence and not the extent of MVO, which is the more important marker for adverse LV function.
LV EF from baseline to 4 months was less pronounced in patients with MVO, as compared to those without. Thus, the presence of MVO, as demonstrated by a CMR study during the first few days after STEMI, is a marker of an unfavorable recovery over time of jeopardized myocardium. MVO was also associated with a large final infarct size, and this association was still highly significant after adjustment for other markers of outcome, including myocardium at risk. Of note, none of our patients had collateral flow to IRA and, except for 3 patients with TIMI 2-flow, all patients in our study had normalization of macrovascular flow after PCI. Our results confirm and extend previous reports on the association between MVO and variables of LV function after PCI-treated STEMI. In patients with presence of MVO, we also found change in LV ESV between baseline and 4 months than in patients without MVO. However, in contrast to other reports, after adjustment for confounders, the change in LV EDV was not significantly different in the 2 groups. Other baseline variables, including myocardium at risk, were of importance for the remodeling process, as expressed by a significant association with LVEDV. This observation corresponds with the results of Lund and coworkers, concluding that infarct size measured by early CMR is a stronger predictor of remodeling than MVO. Some investigators have calculated the size and extent of MVO and related this to LV outcome variables. However, according to Nijveldt and co-workers, it is the presence and not the extent of MVO, which is the more important marker for adverse LV function.