Swelling processes of underground plant storage organs have been extensively studied in tuber as compared with other underground stems. Great changes have been found in genetic and morphometric processes during the formation of storage organs. Similar with tuber development of potato, corm formation of Sagittaria trifolia is also under a strict photoperiodic control. The formation of corm is promoted in short days, but prolongs in long days. It is reported that the signal of photoperiodic control is perceived by the leaf, and then transported via the phloem to the vegetative shoot apex or underground stolon tips, which promotes the transition of storage organ. Flowering Locus T, LKP2, CONSTANS and GIGANTEA has been found to be involved in the signal Chlorhexidine hydrochloride transduction of photoperiodic control, and these gene expressions affect the formation of storage organ. Chen et al. find that formation of storage organ is promoted by StBEL5 and KNOX through repressing the gibberellin StGA20ox1 biosynthesis under SD condition. At the same time, the expression of StBEL5 is enhanced by miR172, suggesting that long distance transport of RNA signal also participates in the formation of underground storage organ. In addition, PHYB is also involved in the formation of storage organ in SD. Decreasing the levels of PHYB in transgenic plants lead to the formation of storage organ both in SD and LD. Compared with transgenic plants, control can form storage organ in SD, suggesting that plants lose the inhibitory effect on tuberization caused by LD. At present, high sucrose content is reported to be the optimal condition required for the formation of storage organs. During the early stages of storage organ development, it requires an active sucrose transporter to trigger the formation of storage organ, which indicates that the role of sucrose is necessary for the ![]() formation of storage organ at the initial swelling stages. Evidence shows that several phytohormones including: gibberellic acid, cytokinin, jasmonic acid, abscisic acid, indole acietic acid, ethylene and jasmonate are also involved in the initiation and regulation of growth in these storage organs. It has been reported that exogenous application of GA acts as an inhibitor of tuber induction. Overexpression of GA oxidase gene in transgenic potato plants postpones the tuber development. Whereas, inhibition of this gene results in an early tuberization than wild type plants. Cytokinin and jasmonic acid promote the tuber induction and elongation. Bhat et al found that exogenous cytokinin is necessary to induce formation of tuber in ginger due to improvement of photosynthesis. ABA shows high correlation to tuber formation because ABAdeficient potato plants show retarded tuberization. Exogenous application of auxin on the decapitated peas and potatoes inhibits the formation of axillary buds. Ethylene, produced by almost all plants mediates a variety of developmental processes in plants, such as seed germination, lateral bud stimulation, adventitious rooting, overcoming dormancy and organ senescence and abscission. Exogenous ethylene is believed as an inducer for the tuberization in potato and root bulking in carrots. Just like the other storage organs, corm of Sagittaria trifolia is also an 4-(Benzyloxy)phenol important edible product, and the developmental processes of this kind of storage organ is regulated by many genes. These differentially expressed genes promote the formation of corm.
formation of storage organ at the initial swelling stages. Evidence shows that several phytohormones including: gibberellic acid, cytokinin, jasmonic acid, abscisic acid, indole acietic acid, ethylene and jasmonate are also involved in the initiation and regulation of growth in these storage organs. It has been reported that exogenous application of GA acts as an inhibitor of tuber induction. Overexpression of GA oxidase gene in transgenic potato plants postpones the tuber development. Whereas, inhibition of this gene results in an early tuberization than wild type plants. Cytokinin and jasmonic acid promote the tuber induction and elongation. Bhat et al found that exogenous cytokinin is necessary to induce formation of tuber in ginger due to improvement of photosynthesis. ABA shows high correlation to tuber formation because ABAdeficient potato plants show retarded tuberization. Exogenous application of auxin on the decapitated peas and potatoes inhibits the formation of axillary buds. Ethylene, produced by almost all plants mediates a variety of developmental processes in plants, such as seed germination, lateral bud stimulation, adventitious rooting, overcoming dormancy and organ senescence and abscission. Exogenous ethylene is believed as an inducer for the tuberization in potato and root bulking in carrots. Just like the other storage organs, corm of Sagittaria trifolia is also an 4-(Benzyloxy)phenol important edible product, and the developmental processes of this kind of storage organ is regulated by many genes. These differentially expressed genes promote the formation of corm.
Essentially all above processes are controlled by the signal molecules and environmental factors
Leave a reply
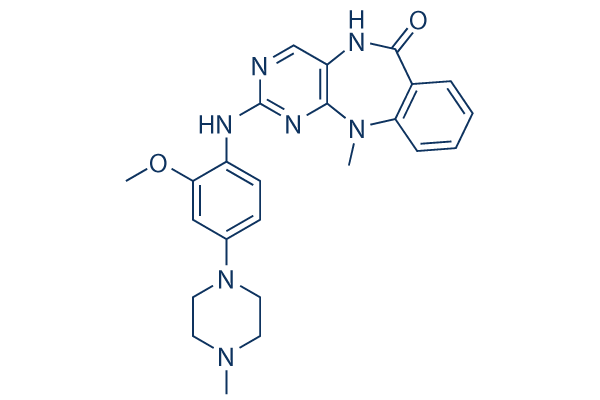 cooperates with Myc to promote cell proliferation. Recently, YAP has been shown to play a critical role in stem cell biology. It is induced during pluripotent stem cell reprogramming, whilst silencing of YAP reduces the pluripotency of embryonic stem cells. YAP promotes ovarian cancer progression, and high levels of nuclear expression are inversely associated with patient survival. In particular, YAP is associated with clear cell ovarian tumors, an ovarian malignancy subtype with poor prognosis. YAP has also been shown to play an oncogenic role in esophageal squamous cell carcinoma. In liver cancer, microRNA-mediated inhibition of YAP inhibits tumor characteristics including cell proliferation and invasion. Conversely, there are reports showing an opposite, tumor suppressive, role of YAP in promoting p73-mediated apoptosis. In breast and head and neck cancers, YAP has been shown to act as a tumor suppressor in certain circumstances. TAZ is structurally homologous to YAP, is likewise inhibited by the Hippo pathway, and also promotes EMT-mediated cancer progression. TAZ regulates mesenchymal stem cell differentiation by modulating Runx2- and PPAR gamma-dependent gene expression.
cooperates with Myc to promote cell proliferation. Recently, YAP has been shown to play a critical role in stem cell biology. It is induced during pluripotent stem cell reprogramming, whilst silencing of YAP reduces the pluripotency of embryonic stem cells. YAP promotes ovarian cancer progression, and high levels of nuclear expression are inversely associated with patient survival. In particular, YAP is associated with clear cell ovarian tumors, an ovarian malignancy subtype with poor prognosis. YAP has also been shown to play an oncogenic role in esophageal squamous cell carcinoma. In liver cancer, microRNA-mediated inhibition of YAP inhibits tumor characteristics including cell proliferation and invasion. Conversely, there are reports showing an opposite, tumor suppressive, role of YAP in promoting p73-mediated apoptosis. In breast and head and neck cancers, YAP has been shown to act as a tumor suppressor in certain circumstances. TAZ is structurally homologous to YAP, is likewise inhibited by the Hippo pathway, and also promotes EMT-mediated cancer progression. TAZ regulates mesenchymal stem cell differentiation by modulating Runx2- and PPAR gamma-dependent gene expression.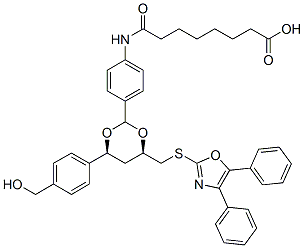 intranasally immunized with NTHi OMVs showed significant reductions in their nasopharyngeal
intranasally immunized with NTHi OMVs showed significant reductions in their nasopharyngeal