In EC by activating NFkB thus promoting pro-inflammatory 3,4,5-Trimethoxyphenylacetic acid angiogenesis, which has been associated with increased adventitial neovascularization and IH. IL-8 also leads to proliferation and migration 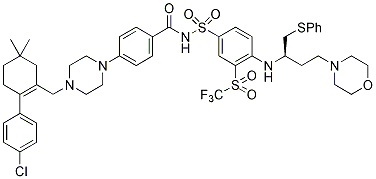 of vascular SMC thereby Chlorhexidine hydrochloride contributing to IH. IL-6 is a pro-inflammatory cytokine mainly secreted by activated macrophages and lymphocytes but also by EC and SMC. IL-6 is involved in immune regulation, hematopoiesis, inflammation and oncogenesis. Although little is known about the role of IL-6 in the pathophysiology of IH leading to vein bypass graft failure, several studies have demonstrated that IL-6 is pro-atherogenic through promoting EC dysfunction, SMC proliferation and migration as well as recruitment and activation of inflammatory cells. Besides IL-8 and IL-6 several other genes from these pathways have been associated with vascular remodeling and hence could affect vein graft implantation injury. In particular, we noted increased levels of transcriptional regulators such as NF-kB, cytokines and cytokine receptors such as IL1A, IL1B and IL1R2, regulators of extracellular matrix such as MMP2, COL1A1 and collagen type 1; and decreased levels of regulators of cell differentiation such as serum response factor. Based on the backpropagation networks, we also delineated the top focus gene hubs that had the greatest interaction density in both EC and SMC. The choice of these focus gene hubs was based on the fact that they provide maximum stability to the backpropagation network. In fact, targeting any of these genes as modeled by their removal from the network offers the most effective means to disrupt the network. Among those focus gene hubs, IL-6, INSR and IGF1R are the genes showing the greatest interaction density, closely followed by IL-8, IL-15 and FGFR2. In fact, among those genes, IL-6 and IL-8 were the most up-regulated genes, whereas INSR, IGF1R and FGFR2 were down-regulated. These results further validate the key role of IL-6 and IL-8 as pathogenic, and therefore as high profile therapeutic targets to prevent vein graft implantation injury. We have validated the up-regulation of IL-6 and IL-8 in graft EC and SMC by qRT-PCR, and confirmed that IL-6 significantly increases from 2 H and up to 7 D post-implantation while IL-8 is up-regulated at all time-points including the 30 D time-point in both cell types. Using qRT-PCR we also validated the upregulation of Coll11A1 at the later time-points, 7 and 30 D suggesting a potential role for extracellular remodeling in driving the healing process, while being the major component in lesion of implantation injury. Current work in our laboratory is aimed at developing local siRNA based therapies to concomitantly target IL-6 and IL-8 secretion within the vein graft and evaluate how this would impact vein graft implantation injury. In conclusion, this study represents the first comprehensive analysis of the genomic response to vein graft implantation injury in a large animal model. LCM has made it possible to separately define the genomic response of EC from that of medial SMC. Our data indicates that a robust genomic response begins by 2 H, peaks at 12�C24 H, starts resolving by 7 D, and declines markedly by 30 D. Inflammatory pathways dominate the early response, followed by modulators of cell cycling, and culminate in pathways involved in extra-cellular matrix remodeling. By using a back-propagation based systems biology analysis of the data, we were able to establish a temporal and causative link between these pathways that helped us identify the molecular signature of vein graft implantation injury, including high intensity hubs.
of vascular SMC thereby Chlorhexidine hydrochloride contributing to IH. IL-6 is a pro-inflammatory cytokine mainly secreted by activated macrophages and lymphocytes but also by EC and SMC. IL-6 is involved in immune regulation, hematopoiesis, inflammation and oncogenesis. Although little is known about the role of IL-6 in the pathophysiology of IH leading to vein bypass graft failure, several studies have demonstrated that IL-6 is pro-atherogenic through promoting EC dysfunction, SMC proliferation and migration as well as recruitment and activation of inflammatory cells. Besides IL-8 and IL-6 several other genes from these pathways have been associated with vascular remodeling and hence could affect vein graft implantation injury. In particular, we noted increased levels of transcriptional regulators such as NF-kB, cytokines and cytokine receptors such as IL1A, IL1B and IL1R2, regulators of extracellular matrix such as MMP2, COL1A1 and collagen type 1; and decreased levels of regulators of cell differentiation such as serum response factor. Based on the backpropagation networks, we also delineated the top focus gene hubs that had the greatest interaction density in both EC and SMC. The choice of these focus gene hubs was based on the fact that they provide maximum stability to the backpropagation network. In fact, targeting any of these genes as modeled by their removal from the network offers the most effective means to disrupt the network. Among those focus gene hubs, IL-6, INSR and IGF1R are the genes showing the greatest interaction density, closely followed by IL-8, IL-15 and FGFR2. In fact, among those genes, IL-6 and IL-8 were the most up-regulated genes, whereas INSR, IGF1R and FGFR2 were down-regulated. These results further validate the key role of IL-6 and IL-8 as pathogenic, and therefore as high profile therapeutic targets to prevent vein graft implantation injury. We have validated the up-regulation of IL-6 and IL-8 in graft EC and SMC by qRT-PCR, and confirmed that IL-6 significantly increases from 2 H and up to 7 D post-implantation while IL-8 is up-regulated at all time-points including the 30 D time-point in both cell types. Using qRT-PCR we also validated the upregulation of Coll11A1 at the later time-points, 7 and 30 D suggesting a potential role for extracellular remodeling in driving the healing process, while being the major component in lesion of implantation injury. Current work in our laboratory is aimed at developing local siRNA based therapies to concomitantly target IL-6 and IL-8 secretion within the vein graft and evaluate how this would impact vein graft implantation injury. In conclusion, this study represents the first comprehensive analysis of the genomic response to vein graft implantation injury in a large animal model. LCM has made it possible to separately define the genomic response of EC from that of medial SMC. Our data indicates that a robust genomic response begins by 2 H, peaks at 12�C24 H, starts resolving by 7 D, and declines markedly by 30 D. Inflammatory pathways dominate the early response, followed by modulators of cell cycling, and culminate in pathways involved in extra-cellular matrix remodeling. By using a back-propagation based systems biology analysis of the data, we were able to establish a temporal and causative link between these pathways that helped us identify the molecular signature of vein graft implantation injury, including high intensity hubs.
Stimulates vascular endothelial growth factor expression and the autocrine activation of VEGF Receptor
Leave a reply
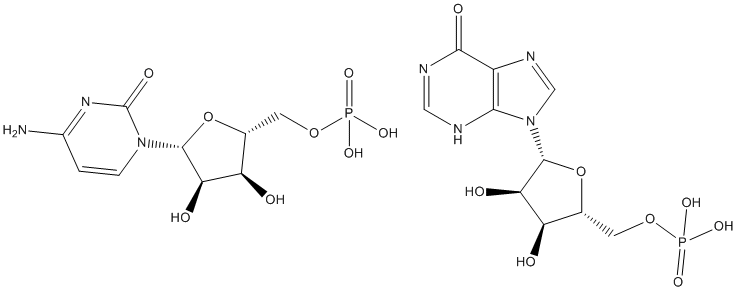 superficial layers �C which include the SO and the retino-recipient layers- produce a decreasing rostro-caudal gradient which extends along the entire tectal axis. This gradient can be appreciated at E5�CE6 �Cwhen the EphA3-positive temporal RGC axons have not yet invaded the tectum- and presents the highest level of expression around E12 when the retinotectal mapping is taking place. The EphA3 expression decreases postnatally when the retinotectal mapping has finished. As axonal ephrin-As are the potential receptors for tectal EphA3, we analyzed the developmental pattern of expression of ephrin-A2 by performing immunocytochemistry on retinal sections coinciding with the naso-temporal axis from E4 to P7.
superficial layers �C which include the SO and the retino-recipient layers- produce a decreasing rostro-caudal gradient which extends along the entire tectal axis. This gradient can be appreciated at E5�CE6 �Cwhen the EphA3-positive temporal RGC axons have not yet invaded the tectum- and presents the highest level of expression around E12 when the retinotectal mapping is taking place. The EphA3 expression decreases postnatally when the retinotectal mapping has finished. As axonal ephrin-As are the potential receptors for tectal EphA3, we analyzed the developmental pattern of expression of ephrin-A2 by performing immunocytochemistry on retinal sections coinciding with the naso-temporal axis from E4 to P7.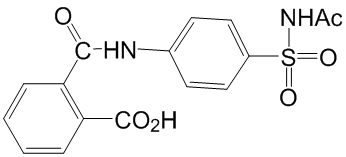 of specific attractant or repellent molecules. However, it is not trivial to identify such specific compounds and further studies should be performed in order to address this point. These unexpected and interesting results strongly suggest that the biological role of the RND-4 and RND-9 efflux pumps might not be restricted to the sole transport of toxic compounds, but also that their function might be
of specific attractant or repellent molecules. However, it is not trivial to identify such specific compounds and further studies should be performed in order to address this point. These unexpected and interesting results strongly suggest that the biological role of the RND-4 and RND-9 efflux pumps might not be restricted to the sole transport of toxic compounds, but also that their function might be 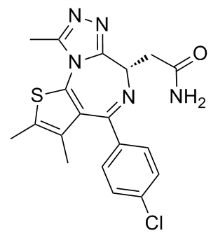 biological specimens, cell, and RNA isolation techniques, hybridization protocols, and statistical methods applied to identify differentially expressed clones.
biological specimens, cell, and RNA isolation techniques, hybridization protocols, and statistical methods applied to identify differentially expressed clones.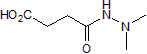 and transcription termination by bacteriophage HK022 Nun protein. A key role for Mfd as an enhancer of UvrA turnover in E. coli cells has also been recently demonstrated. The well characterized Mfd from E. coli is a 130 kDa monomeric protein having modular architecture specialized for different functions. The N-terminal domain shares a high degree of structural homology with UvrB protein of NER pathway. The NTD is known to interact with UvrA protein, which is molecular matchmaker of NER pathway, and this interaction is responsible for enhanced rates of repair. The central portion of Mfd consists of RNA polymerase interacting domain which binds to b subunit of RNA polymerase. The C-terminal domain of Mfd harbors seven signature motifs of super-family 2 helicases including ATPase motifs. In addition, CTD contains a TRG motif required for translocation along the DNA. TRG motif as the name implies, is highly homologous to RecG protein, which is known to be involved in branch migration of Holliday junctions during recombination. Pathogenic bacteria continuously encounter multiple forms of stress in their hostile environments, which leads to DNA damage. Genes involved in DNA repair and recombination may play an important role in the virulence of pathogenic organisms. M. tuberculosis is a gram positive, acid fast bacterium and one of the most formidable human pathogen. DNA repair pathways in mycobacteria appear to be crucial for their survival at different stages of infection. Sequencing of M. tuberculosis genome revealed the presence of NER associated genes including a putative mfd. In this work, we describe the functional characterization of MtbMfd and discuss its unusual properties. This is the first detailed analysis of the biochemical
and transcription termination by bacteriophage HK022 Nun protein. A key role for Mfd as an enhancer of UvrA turnover in E. coli cells has also been recently demonstrated. The well characterized Mfd from E. coli is a 130 kDa monomeric protein having modular architecture specialized for different functions. The N-terminal domain shares a high degree of structural homology with UvrB protein of NER pathway. The NTD is known to interact with UvrA protein, which is molecular matchmaker of NER pathway, and this interaction is responsible for enhanced rates of repair. The central portion of Mfd consists of RNA polymerase interacting domain which binds to b subunit of RNA polymerase. The C-terminal domain of Mfd harbors seven signature motifs of super-family 2 helicases including ATPase motifs. In addition, CTD contains a TRG motif required for translocation along the DNA. TRG motif as the name implies, is highly homologous to RecG protein, which is known to be involved in branch migration of Holliday junctions during recombination. Pathogenic bacteria continuously encounter multiple forms of stress in their hostile environments, which leads to DNA damage. Genes involved in DNA repair and recombination may play an important role in the virulence of pathogenic organisms. M. tuberculosis is a gram positive, acid fast bacterium and one of the most formidable human pathogen. DNA repair pathways in mycobacteria appear to be crucial for their survival at different stages of infection. Sequencing of M. tuberculosis genome revealed the presence of NER associated genes including a putative mfd. In this work, we describe the functional characterization of MtbMfd and discuss its unusual properties. This is the first detailed analysis of the biochemical